Tansgénèse. Exemples chez le maïs (résistance à la pyrale), pomme de terre (résistance au doryphore), soja (résistance au herbicides), terminator des semences تحوير المورثات . أمثلة الذرة و البطاطس و فول الصويا
Tansgénèse. Exemples chez le maïs (résistance à la pyrale), pomme de terre (résistance au doryphore), soja (résistance au herbicides terminator des semences) تحوير المورثات . أمثلة الذرة و البطاطس و فول الصويا
La
transgénèse peut concerner plusieurs espèces de plantes dont le maïs (ذرة, maize ), la pomme de terre ( بطاطس , potato), le soja (فول الصويا ,
soybean) tranformées pour la résistance à la pyrale (lépidoptère), le doryphore (coléoptère) et herbicides, respectivement. La technologie 'terminator' des semences est expliquée
-
Faire un Quiz formatif (Contrôle noté) sur les Biotechnologies
des Plantes et Marqueurs moléculaires
Cas
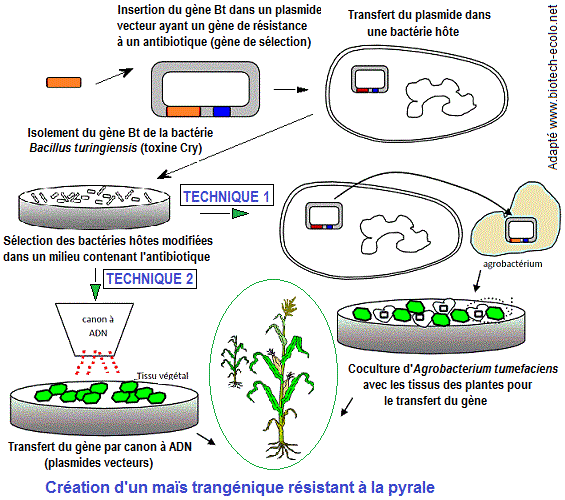
1. Maïs résistant à la chenille de la pyrale (lépidoptères)
ذرة مقاومة لأسروعة الفراشة - حرشفيات
الأجنحة
En
se référant au schéma ci-dessous relatif à
la création d'un maïs transgénique (plante OGM),
repérer les grandes étapes de la transgénèse
couramment suivies dans la mise au point de plantes transgéniques
et concernent :
1. Repérage d'un caractère intéressant dans un organisme vivant et identifier la protéine
2. Identification et isolement du gène d'intérêt
3. Réalisation et amplification d'une construction génique
4. Transfert de l'ADN = Introduction d'un ADN étranger dans une cellule de Maïs (différentes méthodes)
5. Contrôle de l'efficacité du transfert chez l'hôte
6. Sélection des cellules exprimant le gène ajouté par tri
Quels
sont les deux modes de transfert du transgène de la résistance du Maïs à la chenille de la pyrale?
Pourquoi la toxine Cry1Ab n'est pas toxique pour l'Homme?
Des
variétés de maïs ont été transformées par des firmes privées pour produire dans leurs tissus la toxine Cry1Ab de Bacillus thuringiensis (Bt), active contre
la pyrale du maïs.
Cette stratégie de lutte offre plusieurs avantages :
- la toxine Cry1Ab n'est active que sur
les insectes , aucune toxicité n'a été
mise en évidence, ni pour les animaux domestiques ni pour
l'homme.
- la toxine est produite principalement
dans les parties vertes de la plante , qui ne sont jamais
consommées par l'homme ;
- les premiers essais ont montré une remarquable efficacité
de ces maïs
Vidéo
sur la toxine Cry du Bacillus thuringiensis (Bt) et la transformation
génétique des plantes pour la résistances aux
insectes (narration Ar):
Cas 2. Pomme de terre résistance au doryphore (coléoptère) بطاطس مقاوم
للخنفساء - مغمدات الأجنحة
Importance
de la culture de la pomme de terre au Maroc. Au Maroc,
la pomme de terre (potato, بطاطس) (Solanum tuberosum)
est la première culture maraîchère des points
de vue superficie et production (25% des cultures maraîchères).
La production atteint annuellement 1,4 million de tonnes. Celle-ci
est destinée principelement pour le marché local (les marocains sont des gros consommateurs des pommes de terre).
Au
Maroc , les insectes ravageurs causent des dégâts
pour la culture de la pomme de terre. C'est le cas de la noctuelle
terricole Hydraecia micacea (ou ver gris, noctuelle
mineuse de la pomme de terre), appelée aussi en Amérique
du Nord 'Perce-tige de la pomme de terre', est un insecte
de l'ordre des lépidoptères: Sur feuilles, les
jeunes chenilles dévorent le parenchyme des feuilles.
Il ne reste que l'épiderme desséché.
Quand l'attaque est avancée, la culture semble grillée.
Sur tubercule: les attaques de chenilles laissent des galeries
qui évoluent en pourriture. Le Traitement au début
de l'infestation avec des insecticides de contact à
base de méthamidophos, méthomyl, chlopyriphos
et de parathion. Y'a-t-il besoin
de créer des pommes de terre transgéniques résistantes
à la noctuelle terricole au Maroc ?
Extension de l'agriculture dans des conditions difficiles où la pomme de terre peut pousser. Au coeur du Sahara marocain, à plus de 1000 km au sud d'Agadir et à 350 km
des frontières mauritaniennes, une pointe de verdure
surgit du désert. Il est difficile d'en croire ses
yeux. Sur 25 ha, s'étend une exploitation agricole
de pointe. ..en raison de la qualité du climat à
Dakhla, plusieurs autres variétés peuvent y
être exploitées. Ainsi, pour la production sous
serre sont cités, outre les melons et les pastèques,
le poivron, l'aubergine, le concombre, la banane, l'ananas
et le raisin de table. S'ajoute à cela la production
de plein champ (pommes de terre, oignon, ail, concombre, courgette,
artichaut, luzerne, maïs, banane).. Lire
la suite.
Le doryphore (Leptinotarsa decemlineata, coléoptère, مغمدات الأجنحة, Coleoptera, Beetles)
Le doryphore est le plus important ravageur de la pomme de terre au Canada et dans plusieurs régions aux états-Unis. Il se nourrit de plantes de la famille des Solanaceae. Parmi cette famille, on retrouve la pomme de terre, la tomate et l'aubergine qui sont sensibles
à ses attaques. Menace: En Afrique, le doryphore a été signalé
en Libye, mais le Maroc en est indemne (Alain Fraval,
2001. Le doryphore : un grand conquérant fatigué
?, OPIE), toutefois sa présence en Libye n'était
pas confirmée en 2006. Son
potentiel d'expansion est très important : il pourrait
s'étendre dans toutes les régions tempérées
du globe où la pomme de terre est cultivée
et où il est encore absent, notamment en Asie orientale,
dans le sous-continent indien, en Afrique du Nord et en
Afrique australe, en Amérique latine, en Australie
et en Nouvelle-Zélande. Le réchauffement
climatique pourrait favoriser son extension vers le
nord.
De nombreux insecticides sont utilisés pour tenter d'éliminer
le doryphore, mais ils sont peu efficaces contre les pucerons
se déplaçant rapidement d'un endroit à
un autre, véhiculant le PLRV.
Depuis
quelques années, des variétés de
pomme de terre modifiées génétiquement
pour être résistantes au doryphore sont
disponibles. Un gène de la bactérie Bacillus
thuringienssis (BT) a été introduit
dans les plants de pomme de terre.
Bactérie Bacillus turingiensis avec la toxine Cry
La plante peut donc produire elle-même la toxine
du BT qui détruit le système digestif des larves
et des adultes du doryphore et provoque ainsi leur mort.
La culture de variétés de pommes de terre génétiquement
modifiées (GM) a été approuvée
dans le continent Nord Américain et autorisée
depuis 1994 aux USA et 1995 au Canada, avec la résistance
au doryphore seule ou une résistance
associée avec une résistance au potyvirus (PVY)
ou au luteovirus (PLRV) depuis 1998 aux USA et
1999 au Canada. Ces
variétés OGM sont exportées, depuis 2001,
au Japon et en Australie où elles sont autorisées
en alimentation humaine et animale.
En
génie génétique, les tests d'efficacité
de différentes constructions géniques
ont été éssayés au champ. La pomme
de terre est sensible au Colorado Potato Beetle (doryphore)
et au puceron vert du pêcher qui transmet le
PLRV provoquant l'enroulement des feuilles et la nécrose
des tubercules.
Se
déplaçant rapidement, le doryphore échape aux
insecticides. L'obtention de plantes transgéniques permettant
l'expression conjointe du gène Cry3A (codant une toxine
de Bt efficace sur doryphore) et du gène capside du PLRV
est décrite comme permettant une excellente protection.
Néanmoins,
le doryphore peut devenir résistant au BT rapidement si on
emploie les variétés OGM de façon abusive sans
utilisation d'autres moyens de lutte.
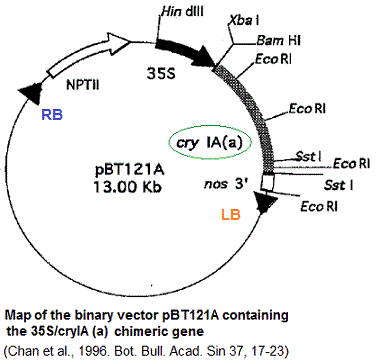
Le
gène cryIA(a) de taille 2 kb, a été
isolé du DNA du plasmide de la bactérie Bacillus
turingiensis (Bt) par PCR (Schnepf et al., 1985). Deux séquences
qui serviront d'amorces (amorce 5' et amorce 3', voir ci dessous)
pour la PCR, ont élé isolées de la région
codante du gène. Elles serviront pour amplifier le fragment
de 2 kb.
le
gène néomycine phosphotransférase II (NPTII)
porté par le
vecteur binaire a été utilisé pour la sélection
des transformants (plantes) mis en culture dans un milieu contenant
la kanamycine. Le gène nopaline synthase (nos) = promoteur,
terminateur
1/
Par quelle méthode les auteurs ont fait le transfert du transgène?
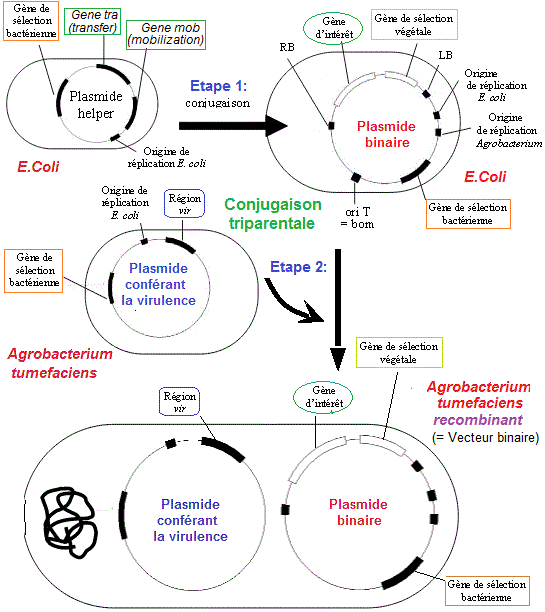
Il
s'agit d'un transfert
binaire. Ici les plants de pomme de terre ont été
inoculés avec Agrobacterium tumefaciens C58C1 (= plasmide
hlper (pGV2260) + plasmide pBT121A). Le plasmide Helper va stimuler
l'introduction du plasmide recombiné dans A. tumefasciens
qui se retrouve avec deux plasmides.
2/
Comment les auteurs révèlent la présence du
DNA du transgène intégré dans la plante hôte?
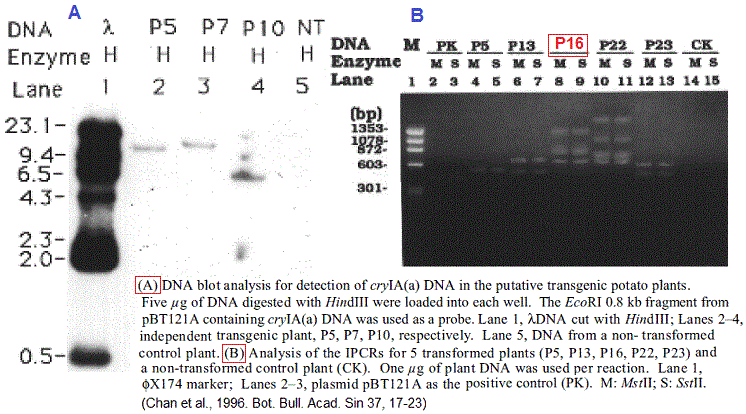
Préparation
d'une sonde radioactive à partir du transgène
cryIA/ Par hydrolyse avec E. coli du transgène,
un fragment d'environ 0,8 kb du gène cryIA, a été
isolé et marqué au 32P radioactif. Cette
sonde s'hybridera avec l'ADN extrait des feuilles des transformants
(plantes) et préalablement digéré par HindIII
et séparé ensuite par électrophorèse
sur gel. Ainsi cette méthode 'southern blotting ' a
permis de révéler des fragments d'ADN de 11 kb, 14
kb et 6,5 kb pour les plantes P5, P7 et P10, respectivement. Cela
démontre l'intégration du gène cryIA(a) dans
le génome des transformants (voir figure). Voir caractérisation
moléculaire des transgènes (cours).
Share this information on social networks تبادل المعلومة
عبر المواقع الإجتماعية
Analyse
d'article (suite). Question 3.
3/ Comment les auteurs se sont rendus compte du site d'insertion du
transgène et du nombre de copies du intégrées
dans l'ADN de chaque plante de pomme de terre?
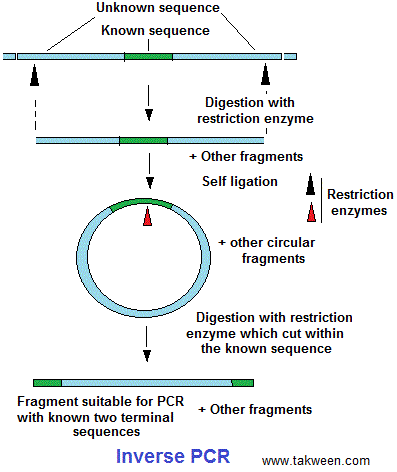
Ils ont procédé par une technique appelée 'inverse PCR' (IPCR) . Comme le montre la figure B beaucoup de plants ont intégré le transgène en plusieurs copies (1-4). La PCR inverse est une variante de la PCR utilisée quand on connaît uniquement une seule séquence (ex. un transgène) entourée par des séquences inconnues et où on ne dispose pas d'amorces pour l'amplification (voir figure). On
procède par:
- Digestion sur les séquences inconnues entourant la séquence connue (transgène).
- Conversion des fragments d'ADN obtenus en ADN circulaire par une ligation interne.
- Digestion des ADN circulaires par une enzyme de restriction qui opère uniquement au niveau de la séquence
connue (transgène).
On en génère des fragments d'ADN circulaire qu'on peut amplifier par des amorces qui reconnaissent une partie
de la séquence connue (transgène).
Avec la PCR inverse; on peut se rendre compte où le transgène s'est inséré. En effet, l'inconvénient de la PCR conventionnelle et l'exigence de deux amorces devant être fixées aux extrémités du gène à amplifier. Avec un gène connu positionné à côté du gène inconnu, on peut créer à partir du premier ces deux amorces pour amplifier le gène inconnu.
Inverse
PCR is especially useful for the determination of insert locations. For example, various retroviruses and transposons randomly integrate into genomic DNA. To identify the sites where they have entered,
the known, "internal" viral or transposon sequences can
be used to design primers that will amplify a small portion of the
flanking, "external" genomic DNA. The amplified product
can then be sequenced and compared with DNA databases to locate
the sequence which has been disrupted. (Wikipedia)
4/ Comment expliquer l'absence d'amplification du gène dans 5
plants (parmi 29) ayant survécu dans un milieu de culture contenant
la Kanamycine (propriété due au gène marqueur NPTII)?. Le marqueur est présent, tandis que le gène
d'intérêt est absent !. Le fait que les amorces préparées pour amplifier le transgène CryIA(a) n'ont pas amplié ce gène dans les transformants
peut être expliqué par l'existence de problèmes liés à la réplication et la réparation des transgènes dans les cellules?
5/ Comment les auteurs ont vérifié l'expression du transgène CryIA(a) intégré dans l'ADN de chaque plante de pomme de terre (transcription du gène)?
Les auteurs ont utilisé la technique du 'Northern blotting' qui consiste détecter l'ARN (après transfert
sur membrane) par hybridation avec une sonde marquée
(ici ADN du transgène). Pour analyser l'expression
du transgène CryIA(a) dans les plants transgéniques
de la pomme de terre, l'ARN total a été isolé à partir des feuilles et hybridé avec l'ADN de la région codante du transgène. Les résultas montrent la présence de transcrits du gène dans les feuilles avec des niveaux d'expression différents (voir figure). D'autre part, la révélation par bromure d'éthidium de l'ARN ribosomique (rRNA) démontre que la quantité du RNA total appliquée est approximativement la même pour toutes les plantes transgéniques.
Certains plants ayant intégré plusieurs copies du transgène comme le plant P16 , montrent l'absence de transcription du gène.
6/ Comment les auteurs ont pu vérifier la traduction du transgène dans les plants de pomme de terre?
Ils ont vérifier la traduction du transgène par les tests biochimiques utilisant la technique dot blot
similaire à la technique 'western blotting'. Voir partie du cours relative à la caractérisation
biochimique des transformants. 7/ Comment expliquer l'activité faible de l'expression du transgène (voir figure) dans certains transformants qui contiennent plusieurs copies du transgène?
L'activité
faible de l'expression du transgène CryIA(a) peut être liée à plusieurs facteurs dont
1/ Phénomène de 'silencing' épigénétique
de l'expression du gène par des séquences répétées
de DNA, 2/ Co-suppression par interférence de brins
d'ARN et 3/ DNA méthylation.
Aussi, l'activité faible du transgène peut être
due à l'instabilité du mRNA.
Cas
3. Soja résistant aux herbicides. فول الصويا مقاوم لمبيدات الأعشاب
La
variété de soja commercialisée sous le nom MON 89789 est une variété transgénique
tolérant le glyphosate.
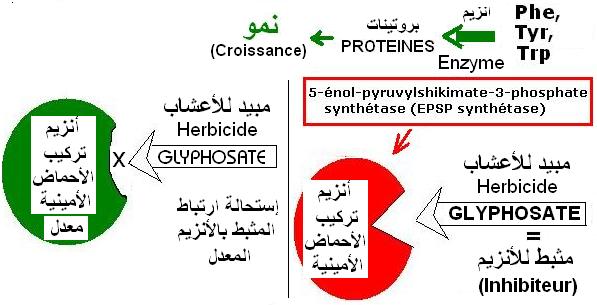
Le
glyphosate est la molécule active d'un herbicide total c'est-à-dire non sélectif utilisé pour le désherbage.
Par
conséquent, il ne permet pas aux agriculteurs de l'utiliser
dans le cadre des traitements qu'ils appliquent aux plantes cultivées
dont le soja.
Le glyphosate est une molécule qui inhibe l'activité d'une enzyme, l'EPSPS (5-EnolylPyruvyl-Shikimate 3-Phosphate Synthase) (Enzyme sensible (ES)) qui dans le chloroplaste catalyse la formation d'acides aminés aromatiques, le tryptophane, la tyrosine et la phénylalanine. La fixation du glyphosate sur l'enzyme est à l'origine de son inhibition.
La synthèse de ces acides aminés étant bloquée, de nombreuses voies métaboliques se trouvent affectées
entraînant en quelques jours la mort du végétal.
Certaines bactéries naturellement présentes dans le sol possèdent un gène codant pour une enzyme EPSPS
(Enzyme tolétante (ET)) tolérant le glyphosate tout
en conservant sa capacité à catalyser la synthèse d'acides aminés.
La variété MON 89789 a été obtenue par modification génétique de la variété commerciale de soja A3224 selon une transformation biologique par Agrobacterium tumefaciens.
Une technique permettant de localiser les gènes sur l'ADN
a montré l'insertion d'une copie du gène cp4-epsps
à un seul locus dans le génome du soja.
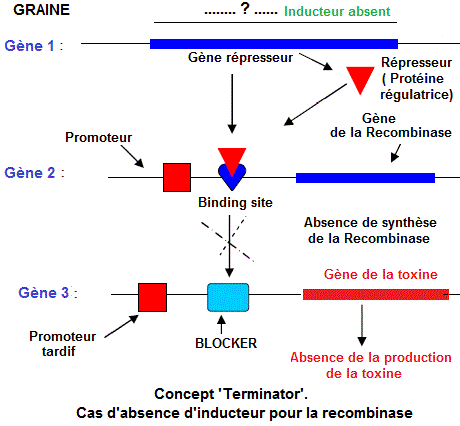
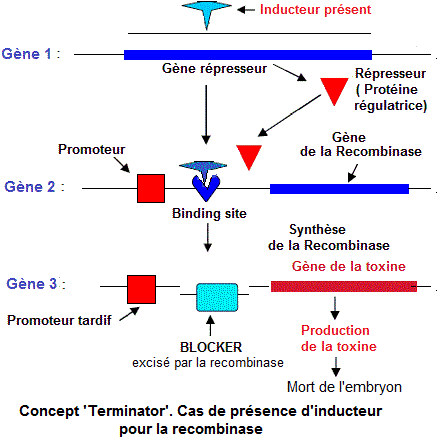
Cas du 'terminator' chez les semences
Le
concept 'Terminator', dont le brevet est détenu par la compagnie
Monsanto, désigne une technique visant l'introduction d'un
transgène (gène exogène
tueur) qui bloque le développement du germe d'un grain.
La plante se développant hibituellement et donnant une récolte normale, produit cependant un grain stérile .
La technologie 'Terminatorr' fonctionne avec trois gènes et un inducteur chimique:
-
Le gène 1 est un gène répresseur
codant pour une protéine régulatrice qui se fixe sur un site opérateur en amont des gènes
de structure et inhibe leur transcription. La protéine
régulatrice du gène 1 (réprésseur) se fixe sur le site de fixation ('binding site' du gène 2
- Le gène 2 ou gène
de l'enzyme recombinase est placé sous le contrôle d'un promoteur. Le site de fixation du répresseur est localisé entre le Promoteur et le gène de la recombinase. Le gène 2 produit une enzyme de restriction qui coupe l'ADN 'blocker' du gène 3.
Les recombinases sont des enzymes permettant la recombinaison génétique. Les DNA recombinases permettent de manipuler la structure du génome et contrôler
l'expression des gènes. Ces enzymes catalysent les réactions d'échange d'ADN dans un sens défini, entre des sites spécifiques déterminés
par de courtes séquences (30 à 40 nucléotides)
propres à chaque recombinase. Ces réactions rendent possibles quatre modules fonctionnels: 1/ Excision/insertion,
2/ Inversion, 3/ Translocation et Echange de cassettes.
- Le gène 3 ou gène
de la toxine létale pour l'embryon . Il est
sous le contrôle d'un promoteur tardif qui est activé
seulement durant le développement de la graine, pendant
la croissance de l'embryon. L'ADN 'Blocker' est localisé
entre le promoteur tardif ('late promoter ') et le gène
3. L'excision de L'ADN 'Blocker' favorise, la transcription
et l'expression du gène de la toxine létale.
Une toxine, ainsi produite, tue l'embryon et rend stériles
les nouvelles semences (pour une reproduction).
- Afin d'activer les gènes 2 (recombinase) et le gène
3 (toxine), un 'inducteur chimique
(tétracycline , par exemple) est répandu
sur les semis par les compagnies de semences comme Monsanto
et Syngeta. Une fois ces gènes activés dans
les semences, on peut avoir la production mais pas le reproduction. Etapes d'activation du gène terminator dans les semences de vente aux agriculteurs:
1/ Introduction de l'inducteur (par exemple du tétracycline ) dans les semences,
2/ L'inducteur bloque le 'binding site' au niveau du gène
2 (recombinase) et empêche la ficxation du répresseur,
3/ L'ARN polymérase se lie au promoteur et commence la transcription
du gène de la recombinase (isomérase) en ARN d'où synthèse de l'enzyme recombinase.
4/ La recombinase taille et coupe la séquence 'blocker',
5/ L'ARN polymérase se lie au 'late promoter ' et transcrit le gène de la toxine.
Il en résulte la synthèse de la toxine qui tue les embryons des graines avant les récoltes.






-northern-blot.gif)
-dot-blot.gif)


{kind=link}