Sélection Assistée par Marqueurs (MAS, Marker Assisted Selection)
La
Sélection Assisteé par Marqueurs ou MAS (Marker Assisted Selection) est basée sur la possibilité de détecter la présence d'un gène ou d'un trait agronomique intéressant par la recherche du marqueur qui lui est étroitement lié (linkage).
Les avantages liés à l'application des marqueurs moléculaires en sélection sont nombreux:
- La MAS est non destructive
- La MAS nécessite peu de tissu végétal
- La MAS n'est pas influencée par des facteurs environnementaux.
Ce type de sélection est particulièrement avantageux lorsque
le caractère étudié est difficile de détection,
coûteux à évaluer ou influencé par les conditions
climatiques ou édaphiques. Avec la SAM, le sélectionneur
peux anticiper la résolution de problèmes liés
à l'adaptation des plantes à de nouvelles contraintes, comme
l'amélioration des plantes pour la résistance aux maladies
dans des régions où le pathogène n'existe pas
encore.
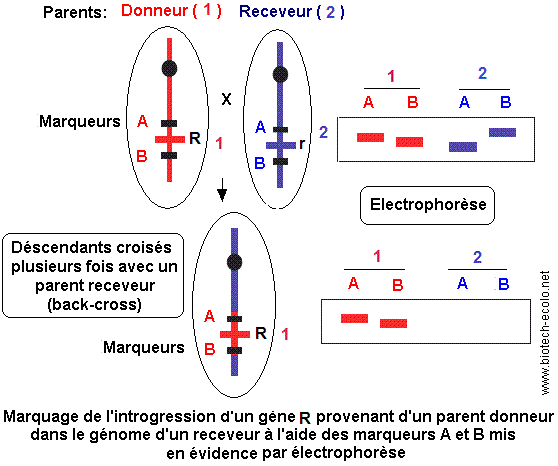
De même, la MAS présente un grand intérêt
dans les programmes d'introgression destinés à
modifier de manière ciblée un matériel génétique
existant, en remplaçant un segment chromosomique initial par un
segment porteur de caractéristiques favorables provenant d'un
autre matériel. Cette approche suppose de croiser deux lignées,
l'une considérée comme le parent donneur (géniteur),
l'autre comme le parent receveur (parent récurrent),
puis d'éliminer progressivement par rétro-croisements
successifs (back-cross) le génome du parent donneur, tout en conservant de façon ciblée le segment (gène) d'intérêt.
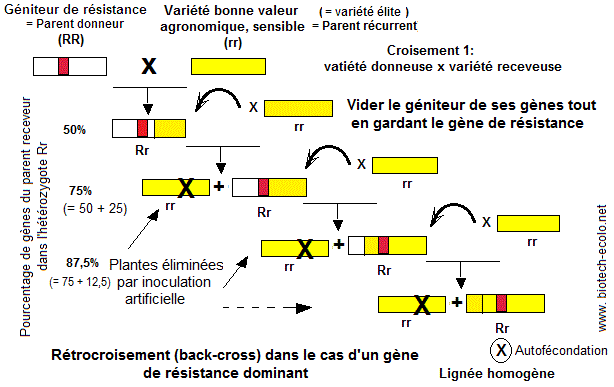
Le rétrocroisement (back-cross) est une méthode d'amélioration pratiquée par les sélectionneurs. Son principe est d'éliminer (vider) progressivement tous les gènes d'un géniteur donné comme un géniteur de résistance (parent donneur), sauf celui qui confère la résistance à une maladie déterminée. Ceci est réalisé par des recroisements successifs de celui ci avec une variété de bonne valeur agronomique (= parent récurrent = parent receveur) . Au cours des recroisements, les gènes du parent récurrent 'remplissent' le géniteur où leur proportion augmente de 50% (première hybridation) à 75% (= 50 + 50/2) (premier recroisement), 87,5%
(= 50 + 75/2) (deuxième), 93,75% (troisième), 96,875% (quatrième recroisement) ,...
D'autre
part, la MAS peut aider à accumuler dans une plante plusieurs gènes de résistance complémentaires pour une même maladie, de façon à obtenir une résistance multigénique, potentiellement plus stable car difficile à contourner par le pathogène. Le phénotype seul ne permet pas de différencier les individus cumulant deux ou plusieurs résistances de ceux qui n'en possèdent qu'une,
rendant le recours aux marqueurs indispensable.
En plus des avantages, la MAS présente des limites dont:
- La MAS n'est compétitive en termes de coût et de temps par
rapport aux méthodes de sélection traditionnelles, lorsque
le phénotype peut être déterminé facilement
(hauteurs des plantes, précocité, résistance à
certaines maladies...)
- La MAS est inefficace pour la sélection de caractères agronomiques
à déterminisme génétique complexe (comme le
rendement, par exemple), gouvernés par un grand nombre de gènes
ou de QTL qui interagissent et dont la plupart sont encore inconnus.
Liens utiles
- Amélioration génétique des plantes. Introduction
- Polyploïdisation, Haploïdisation des plantes
- Modes de reproduction des plantes (autogames, allogames)
- Amélioration génétiques des plantes allogames
- Amélioration génétiques des plantes autogames
- Autogames, allogames. Exercice
- Sélection
des plantes autogames. Exercice
- Sélection
des plantes allogames. Exercice
- RAPD et amélioration de la tomate pour la résistance à la verticilliose, RFLP. Examen S5_2012
- RFLP, RAPD et sélection chez le Cyprès du Japon (examen 2013)
Biotech-ecolo. net. Supports
CHAINE YOUTUBE (abonnement)
Plusieurs vidéos multilingues aux sujets des Biotechnologies et Biochimie

NEWSLETTER